Минулого разу ми зупинилися на обговоренні скорочення скелетних м’язів. Основна ідея полягає в тому, що в руховому нерві виникає потенціал дії. Потім цей потенціал дії передається на м’язову клітину в нейром’язовому синапсі. Він проходить по м’язовій клітині, досягає трубочок, і в результаті мембрана саркоплазматичного ретикулуму стає проникною для кальцію. Кальцій дифундує з саркоплазматичного ретикулуму в рідину клітини, досягає актину і міозину, і за наявності кальцію міозинові поперечні мостики прикріплюються до актину. Потім вони змінюють свою форму і тягнуть актин за собою. За наявності АТФ мостики від’єднуються, перезаряджаються за допомогою енергії АТФ, і, якщо в рідині ще є кальцій, вони прикріплюються до нового активного центру актину, знову втягують його, і процес повторюється. Внаслідок цього м’язові клітини стають дедалі коротшими. Зрештою, коли весь кальцій вичерпується, а точніше, повертається назад у саркоплазматичний ретикулум, активний міозин від’єднується, і м’яз розслабляється.
Таким чином, м’яз залишиться в такому положенні. Наприклад, якщо я напружу свій біцепс і підніму руку, зігнувши її, а потім розслаблю, перестану скорочувати м’язи, рука залишиться зігнутою. Щоб повернути її в початкове положення, мені потрібно скоротити антагоністичний м’яз з іншого боку суглоба, тоді я зможу розтягнути актин і міозин назад у початкове положення. Це за умови, що, наприклад, біцепс підняв руку вгору, і сила тяжіння не опустить руку назад. Інакше мені доведеться використовувати м’язи-антагоністи.
Далі я хотів би зазначити ще кілька моментів. Перша базова одиниця скорочення називається фасцикуляція, або м’язова судома. Якщо перевірити це в лабораторному експерименті, ми беремо м’язову клітину скелетного м’яза і стимулятор. Потім подаємо до м’яза невеликий розряд, стимулюємо потенціал дії, і м’язова клітина різко скорочується. Це відбувається за долю секунди: відбувається викид кальцію, одиничний рух актину і міозину, одиничне всмоктування кальцію в саркоплазматичну мережу і одиничне розслаблення. Тобто клітина один раз смикається і зупиняється. Це одна одиниця м’язового скорочення.
Якщо я захочу більше, мені потрібно запустити новий потенціал дії. Я можу подавати ці потенціали дії в швидкій послідовності, стимулювати безліч клітин раз за разом — тік-тік-тік, робити це швидко-швидко-швидко, і м’язи скорочуватимуться. Якщо робити це досить швидко, м’яз може опинитися в стані постійного скорочення, яке називається тетанус, або оніміння. Якщо скорочувати м’язи дуже швидко і довести їх до стану постійного скорочення, станеться оніміння. Але такий сценарій нам не підходить.
Наприклад, якщо я захочу підняти руку і утримувати її в витягнутому положенні довго, і при цьому стимулюватиму одні й ті самі м’язові клітини знову і знову, вони, зрозуміло, увійдуть у стан тетанічного скорочення. Їх заклинить у напруженому стані надовго, і буквально одразу ви відчуєте такий біль, що мало не здасться. Причина в тому, що кровопостачання до цих клітин, яке забезпечує кисень і поживні речовини та виводить відходи, здійснюється мережею капілярів, що обволікають кожну клітину. Якщо я скорочу цю клітину і залишу її в такому стані, очевидно, що м’язові клітини збільшаться в діаметрі. Це призведе до того, що кровоносні судини поруч із клітиною будуть передавлені, тобто ми перекриємо кровотік до цієї м’язової клітини. Досить швидко клітина вичерпає запаси кисню і палива, внаслідок чого не зможе підтримувати тривале скорочення.
Питання в тому, як нам підтримувати тривале скорочення? Щоб уникнути тетанічного скорочення і оніміння м’язових клітин, ми робимо так, що моторні одиниці скорочуються асинхронно, тобто не в такт одна з одною. Наприклад, якщо я витягнув руку прямо, я можу стояти і тримати її витягнутою цілий ранок, якщо захочу, і не відчуватиму сильного болю. Як я це роблю? Я скорочую кілька моторних одиниць у цій м’язі, потім даю цим моторним одиницям розслабитися, а в цей час скорочуються інші моторні одиниці. Потім даю розслабитися їм і перекладаю роботу на інші. Як сказав мені один студент, таким чином я не спровокую постійне тетанічне скорочення моторної одиниці.
Отже, ми перекидаємо роботу між моторними одиницями, уникаючи тривалого скорочення окремої моторної одиниці. Ми можемо викликати кілька посмикувань однієї моторної одиниці, потім дати їй розслабитися, скоротити інші моторні одиниці, потім треті, і так далі, поки цикл не замкнеться, і ми не почнемо скорочувати їх знову. Фішка в тому, що в кожному окремому м’язовому органі є набагато більше м’язових клітин, ніж ми можемо скоротити за один раз. Дуже рідко ми скорочуємо всі м’язові клітини в окремому м’язовому органі.
Іноді таке трапляється. Час від часу в новинах чути про людину, яка продемонструвала надлюдську силу. Наприклад, якась бабуся бачить, що людину придавило машиною, і раптом вона зсуває цю машину, звільняючи потерпілого. Або хтось піднімає холодильник, щоб звільнити придавленого кота. Якщо я зайду в роздягальню до спортсменів перед важливою грою, видно, як вони намагаються увійти в бойовий настрій. Для чого це? Вони намагаються перекрити механізми, що контролюють ці процеси, щоб скоротити кожну клітину в м’язовому органі одночасно і досягти неймовірних силових показників. Але в новинах не розповідають, який серйозний урон отримує їхня сполучна тканина внаслідок дії такої величезної сили. Потім спортсмен лежить два тижні в ліжку, намагаючись відновитися після свого геройства.
У звичайному режимі наші м’язи працюють не так. Зазвичай м’язові клітини скорочуються не в такт одна з одною, щоб забезпечити тривале скорочення і не пошкодити навколишні тканини. Ми поговоримо про цю тему докладніше і обговоримо інші загальні принципи м’язового скорочення трохи пізніше.
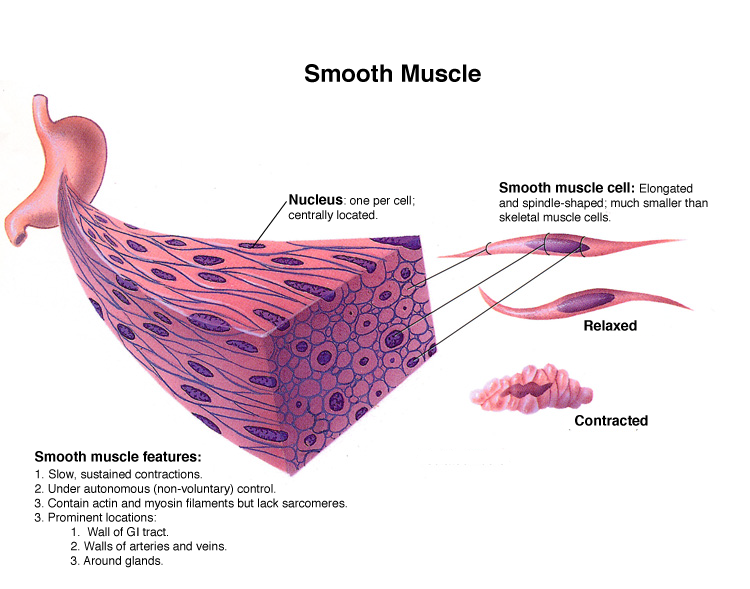
Тож ми обговорюємо три типи м’язів і їхню будову. Скелетні м’язи ми розібрали. Наступний тип м’язів, про який ми поговоримо, — це гладкі м’язи. Спершу подумайте, де ми можемо знайти гладкий м’яз. Вони розташовані в місцях з автоматичним, несвідомим рухом. Ви не можете скоротити гладкий м’яз довільно. Для початку поговоримо про будову самих клітин. Клітина гладкого м’яза — це веретеноподібна клітина з одним ядром у центрі, загострена на кінцях, маленька і коротка. Вона не смугаста, тобто не поперечносмугаста. Коли кажуть, що вони гладкі, мають на увазі саме це — у них немає поперечних смуг. У них також є актин і міозин, але вони розташовані не впорядковано.
Як ми вже визначили, смуги в скелетних м’язах виникають через те, що є маленький диск актину, потім диск міозину, потім знову диск актину і диск міозину, і так далі. Якщо придивитися, як вони перемішані між собою, видно, що навколо кожного міозину розташовані шість актинів. Таке ідеальне впорядковане розташування дозволяє активувати багато міозинових поперечних мостиків одночасно, щоб виробити багато сили за короткий проміжок часу.
Отже, так працюють скелетні, або поперечно-смугасті м’язи. У нас є короткий імпульс сили. У гладких м’язах також є актин і міозин, і вони розташовані вздовж поздовжньої осі клітини, але більш розкидані. Якщо уявити волокна актину і міозину, то фібрили міозину в клітині будуть укладені безладно. Як це впливає на функції гладкого м’яза? По-перше, у нас буде повільне, плавне скорочення.
Якщо взяти клітину гладкого м’яза і простимулювати її, то порівняно з клітиною скелетного м’яза вона працюватиме за тим самим принципом. Подумайте: пропускаємо потенціал дії, виділяємо кальцій, актин і міозин взаємодіють, і за наявності АТФ міозиновий поперечний мостик перезаряджається. Коротше кажучи, механізм той самий, просто актин і міозин упаковані не так щільно і впорядковано.
Візьму клітину гладкого м’яза, дам їй невеликий розряд струмом, запущу процес скорочення, і вона повільно скорочується і повільно розслабляється. Якщо підключити її до динамометра і зобразити графік сили скорочення, то посмикування скелетного м’яза виглядатиме так: ми стимулюємо її, генеруємо скоротливу силу, потім вона розслабляється, і ми отримуємо круту криву. Крива скорочення гладкого м’яза виглядатиме дуже-дуже плавно: вона поступово скоротиться і поступово розслабиться. Це буде дуже повільно і плавно порівняно з сильним різким посмикуванням скелетного м’яза.
Питання, звичайно, чому? Перша причина в тому, що актин і міозин розташовані безладно, не впорядковано. Подумайте ще раз: у скелетних м’язах, як я вже зазначив, спостерігається дуже впорядкована механічна структура актину і міозину. Міозинові поперечні мостики розташовані дуже близько до активних центрів актину, і щойно з’являється кальцій, місток прикріплюється і тягне. У гладких м’язах актин і міозин розташовані деінде, кальцій потрапляє в клітину, і через деякий час міозин знаходить актин, підчіплюється, починає рух і виробляє силу. Волокна не укладені щільно і паралельно, щоб скорочення було миттєвим, як у скелетних м’язах. Нам потрібно розташувати їх для більш плавного скорочення.
Щоб зрозуміти, як виглядають скорочення, уявіть, які рухи ви можете виконувати. Якщо я хочу зрушити з місця, я можу стартувати або рвонути швидко, тому що мої скелетні м’язи скорочуються і розслабляються дуже швидко. Але скорочення гладких м’язів нагадуватиме рухи дощового черв’яка, який повзе по тротуару після дощу. Ви напевно бачили, як черви повзають по доріжках чи калюжах. Черв’як скорочується поступально, повільно скорочуючи один сегмент тіла, потім інший, а потім перший розслабляється. Уявіть, як цей черв’як повзе по доріжці — гладкі м’язи рухаються подібним чином. Рухи черв’яка більше схожі на гладкий тип м’язів, ніж на скелетний.
Отже, перша причина поступального скорочення — це нерівномірне розташування актину і міозину в клітині. Друга причина в тому, що кальцій надходить не з саркоплазматичного ретикулуму. Кальцій дифундує в клітину з позаклітинної рідини. У скелетних м’язах кальцій міститься всередині саркоплазматичного ретикулуму, у цій зеленуватій мережі мембран, яка розташована прямо навколо пучків актину і міозину, її багато по всій клітині. Вона створює величезну площу поверхні для виділення кальцію. Але в клітині гладкого м’яза немає такого густого саркоплазматичного ретикулуму. У нас є лише клітинна мембрана, і кальцій надходить у клітину ззовні. По-перше, відстань для дифузії досить велика, а по-друге, у нас немає великої площі поверхні для ефективної дифузії. Тому скорочення відбувається повільно.
Для розслаблення нам потрібно перекачати кальцій із внутрішньоклітинної рідини у позаклітинну. Знову ж таки, у нас недостатньо площі поверхні, а отже, недостатньо насосів. Тому перекачування кальцію в позаклітинну рідину займе набагато більше часу. У результаті посмикування гладкого м’яза відрізнятиметься. Одиничне скорочення, яке також називається посмикуванням, буде дуже повільним порівняно з швидким посмикуванням скелетного м’яза.
Ще один функціональний момент гладкого м’яза — це так звана спонтанна ритмічність, тобто вбудована. Вона демонструє вбудований ритм. Це означає, що цей тип м’язів не потребує зовнішньої стимуляції руховими нервами. Клітини гладкого м’яза не мають рухових нервів, тобто у них немає нейром’язового синапсу. Вони працюють інакше, мають спонтанний, тобто вбудований ритм скорочення.
Якщо взяти клітину скелетного м’яза, наприклад, біцепса, покласти її на долоню і спостерігати, це буде досить нудне видовище, тому що нічого не відбуватиметься. Ця клітина просто лежатиме на місці і нічого не робитиме. Але якщо взяти клітину гладкого м’яза і спостерігати за нею, не стимулюючи її, то періодично можна побачити, як вона скорочується і розслабляється, потім знову затихає на деякий час, а потім знову скорочується і розслабляється. Нам не потрібно нічого робити, щоб це сталося. У клітині є якийсь вбудований автоматичний механізм, який називається спонтанна ритмічність.
Причина цієї спонтанної ритмічності полягає в коливанні мембранного потенціалу. Він спонтанно коливається вгору-вниз, або, можна сказати, осцилює. Якщо взяти клітину гладкого м’яза, вставити в неї електрод, а інший електрод розмістити зовні, підключити їх до вольтметра і виміряти мембранний потенціал, то в нейроні чи скелетному м’язі, коли ми вставимо електроди, стрілка покаже 70–80 мілівольт, і там залишиться, якщо ми не запустимо потенціал дії. Але в клітині гладкого м’яза мембранний потенціал також буде на рівні 70–80 мілівольт, але з часом він трохи знизиться, потім підніметься, знову знизиться і знову підніметься. Він не буде постійним, а коливатиметься, скакатиме вгору-вниз.
Якщо зобразити це на графіку, ми побачимо, що мембранний потенціал клітини гладкого м’яза варіюватиметься нестабільно. Іноді він падатиме зовсім трохи, іноді трохи більше, вийде нерівномірна амплітудна крива. Як і в будь-якій іншій клітині, її потенціал буде 70–80 мілівольт, але він не буде постійним.
Тепер подумайте, що станеться. Щоб створити потенціал дії, запустити хвилю деполяризації, нам спочатку потрібно досягти порога, тобто зменшити мембранний потенціал до певної точки. Час від часу, чисто випадково, мембранний потенціал падає і досягає порога, запускаючи хвилю деполяризації. Потім мембранний потенціал продовжує свої дрібні коливання, знову перетинає поріг і знову запускає потенціал дії. Хвиля деполяризації працює як будь-який інший потенціал дії: відкриваються потенціалзалежні натрієві канали, і потенціал дії проходить по мембрані клітини, що призводить до притоку кальцію всередину і відповідно до подальшого скорочення.
Нам не потрібно нічого робити з клітинами гладких м’язів — вони спонтанно деполяризуються. Причина в тому, що якщо подивитися на нейрон або клітину скелетного м’яза, то потенціал дії виникає, коли ми знижуємо мембранний потенціал до порогового значення в одному місці. Це провокує відкриття потенціалзалежних каналів. Ми відкриваємо перші потенціалзалежні канали, впускаємо більше натрію всередину, це відкриває наступні потенціалзалежні натрієві канали, впускає нову порцію натрію, і таким чином формується хвиля деполяризації.
У клітинах гладкого м’яза ми бачимо наступне: потенціалзалежні натрієві канали просто відкриваються час від часу самі по собі. Вони чисто спонтанно розкриваються, і я не знаю, як інакше це сказати. Натрієві канали відкриваються, а потім, звісно, знову закриваються. Замість злагодженого механізму, який спрацьовує строго в певний момент, потенціалзалежні канали в клітині гладкого м’яза перебувають у спокої і закриті більшу частину часу. Але періодично, так би мовити, дмухне вітер, і — чпок — канали відкриваються, натрій спрямовується всередину, а потім канали знову закриваються.
Якщо станеться так, що цілий ряд цих каналів відкриється в одному місці в потрібний час, мембранний потенціал у цьому місці впаде до порогового значення і спровокує хвилю деполяризації. Уявіть клітину гладкого м’яза, яка наповнена цими каналами по всій поверхні, і вони відкриваються та закриваються випадковим чином. Але якщо співпаде так, що кілька каналів відкриються в одному місці одночасно, достатня кількість натрію потрапить у клітину, це знизить мембранний потенціал до порогового значення і запустить потенціал дії. Таким чином, тут постійно відбуваються такі безладні коливання через випадкове відкриття цих натрієвих каналів.
Отже, спонтанне скорочення відбувається без будь-яких нервових імпульсів, без будь-яких рухових нервів. Тепер інша ідея, чи радше питання, яке має крутитися у вас на язиці: подумайте про гладкі м’язові клітини, які ви дійсно можете спостерігати. Прийдіть сьогодні додому, подивіться в дзеркало і зверніть увагу на гладкі м’язи райдужки ока. Кольорова частина вашого ока складається з клітин гладкого м’яза, які виходять із зіниці, з отвору в центрі. Їх можна побачити.
Ви коли-небудь уважно розглядали райдужку своїх очей? Візьміть збільшувальне скло і розгляньте очі вашого сусіда по кімнаті чи когось іншого, або знайдіть дзеркало для макіяжу, яке збільшує відображення. Це допоможе вам краще розглянути свої очі. Спостерігайте за своєю райдужкою, стоячи в ванній перед дзеркалом. Зверніть увагу, як вона залишається нерухомою. Ви коли-небудь помічали? Вона трохи розкривається, трохи закривається і весь час перебуває в русі.
Тепер зайдіть у ванну з вимкненим світлом — не в повній темряві, звісно, щоб бачити, куди йдете — і постійте трохи в темряві. Ваші зіниці розширяться. Скільки часу вам потрібно чекати, поки зіниці розширяться? Наприклад, якщо ви заходите в зал кінотеатру посеред дня, скільки вам потрібно, щоб адаптуватися? Коли ви тільки заходите, усе навколо здається в непроглядній темряві, і ви не можете розрізнити номери сидінь. Знайоме, правда? Але якщо постояти хвилину-другу, зіниці розширяться, і ви зможете бачити майже як удень. Але скільки часу потрібно, щоб очі повністю пристосувалися? Ну, три-чотири-п’ять хвилин, мабуть. Постійте у ванній без світла кілька хвилин, поки зіниці повністю не розширяться, а потім різко увімкніть світло і спостерігайте за райдужкою ока. Зіниця швидко звузиться — не миттєво, але досить швидко.
Що ж відбувається? Я щойно сказав, що клітини гладких м’язів не мають рухових нервів. І все ж, коли я вмикаю світло, що однозначно є стимулом, ці м’язові клітини скоротилися і обмежили кількість світла, що проникає, зробивши зіницю меншою. Тож виникає питання: чи не є така реакція прикладом контрольованого процесу? Виходить, клітини гладких м’язів якимось чином знають про яскравість світла? Ні, вони підключені до вашої нервової системи через вегетативні нерви. Що таке вегетативні нерви? Симпатичні та парасимпатичні нерви. Вегетативна нервова система відповідає за регуляцію темпу спонтанної ритмічності.
Давайте подумаємо про це так. Якщо знову уявити наш графік потенціалу, ось тут буде поріг. Ще раз згадайте, що таке поріг. Поріг — це значення мембранного потенціалу, якого потрібно досягти в будь-якому місці клітини гладкого м’яза, щоб запустити потенціал дії. Потенціал дії пробігає по всій поверхні мембрани. Уявіть велику роздуту гладком’язову клітину, і ми бачимо, як по всій мембрані один за одним відкриваються канали. Мембранний потенціал в одному місці клітини трохи відрізняється від інших місць. Але якщо в одному місці станеться так, що багато каналів відкриються, мембранний потенціал досягне порога, і ми отримаємо поширення деполяризації. Тобто відкриємо одні канали, впустимо натрій, потім наступні, ще більше натрію, і так далі — отримаємо хвилю деполяризації.
Ось наш нормальний потенціал спокою, мембранний потенціал коливається так. При викиді вегетативного нейромедіатора — до речі, які вегетативні нейромедіатори ви знаєте? Адреналін у симпатичних нервах і ацетилхолін у парасимпатичних. Якщо я закидаю гладком’язову клітину одним із цих вегетативних нейромедіаторів, ми змістимо коливання мембранного потенціалу ближче до порога. Що станеться? Звісно, він досягне порога раз, другий, третій, четвертий, і ми отримаємо багато потенціалів дії. Коли концентрація нейромедіатора падає, потенціал повертається до звичайного значення.
Якщо я огорну цю клітину іншим нейромедіатором, що, на вашу думку, станеться? Коливальний мембранний потенціал зміститься в інший бік. Який буде результат? Ми взагалі не досягнемо порога. Один нейромедіатор підвищує ймовірність досягнення порога, а інший віддаляє значення мембранного потенціалу далі, знижуючи шанси на перетин порога. Таким чином, нейромедіатори вегетативної нервової системи змінюють співвідношення коливального мембранного потенціалу до порогового значення, підвищуючи або знижуючи ймовірність досягнення мембранним потенціалом цього порога.
У вас виникли питання? Я не прошу вас ставити питання, я просто запитую, який наступний логічний крок вам потрібно знати. Який нейромедіатор за що відповідає? Давайте подумаємо про гладкі м’язи в кровоносних судинах. У стінках кровоносних судин є клітини гладких м’язів. Якщо змусити їх скорочуватися, кровоносна судина стиснеться, а якщо розслабити — розшириться. Тобто клітини гладких м’язів контролюють діаметр кровоносних судин, а разом із цим і кровотік. Якщо розширити кровоносні судини, ми збільшимо кровотік до того місця, куди веде ця судина. Якщо ж простимулювати клітини гладких м’язів до скорочення, підвищити темп їхньої спонтанної ритмічності, вони стиснуть кровоносні судини і зменшать приплив крові.
Спробуймо розібратися. Що дає симпатична стимуляція? Якщо простимулювати симпатичні нерви, виділити адреналін, як це вплине на кровоносні судини? Це змусить їх звузитися в одних місцях і розширитися в інших. Адреналін
Гладкі м’язи в одних місцях реагують на адреналін збільшенням темпу спонтанної ритмічності, а в інших — зниженням темпу спонтанної ритмічності.
Який тип рецептора за що відповідає? Це залежить від місця розташування гладкгої м’яза, хіба не очевидно? У симпатичному стані, у стані «бий або біжи», коли ми займаємося фізичними вправами або, якщо вдаватися до крайнощів, тікаємо від хижака, краще, щоб в одних місцях клітини гладких м’язів розслабилися, а в інших скоротилися. Тому в кожному з цих місць у клітинах є різні рецептори до адреналіну (або епінефрину). Коли ми впливаємо на клітину гладкого м’яза адреналіном, в одних місцях вона скорочується, посилюючи свою спонтанну ритмічність, а в інших розслабляється, тобто послаблює свою спонтанну ритмічність.
Як приклад, один із найпростіших для розуміння — це гладком’язові клітини кровоносних судин. Під час симпатичної стимуляції нам потрібно посилити кровотік до одних ділянок тіла і послабити до інших. І, звісно, реакція гладкого м’яза на адреналін буде протилежною до реакції на ацетилхолін. Подумайте ще раз. Візьмемо кровоносні судини, що постачають кров до вашої шкіри. Коли ці клітини гладких м’язів омиваються адреналіном, це призводить до збільшення швидкості спонтанної ритмічності, що спричиняє стиснення кровоносних судин і збліднення шкіри. Якщо ви щільно поїли і розслаблено сидите в кріслі, ці клітини омиваються ацетилхоліном, що знижує швидкість внутрішньої ритмічності, клітини розслабляються, і кровотік до шкіри збільшується.
Важливо, про що йдеться. Реакція на ці два нейромедіатори буде протилежною. Тобто, якщо адреналін підвищує темп спонтанної ритмічності, то ацетилхолін знижує його, і навпаки. На словах це звучить досить заплутано, але все стає на свої місця, якщо трохи подумати і прокрутити це в голові. Тут не потрібно нічого зазубрювати, просто зрозумійте, як логічно краще організувати цей процес.
Ще раз: ми не кажемо, що вегетативні нейромедіатори адреналін і ацетилхолін викликають скорочення гладких м’язів. Гладка м’яза не потребує зовнішнього стимулу для скорочення. У цьому випадку ми лише регулюємо ймовірність її скорочення шляхом зміни швидкості спонтанної ритмічності, тобто зміни частоти коливань мембранного потенціалу. Якщо вам легше уявляти, що нейромедіатор викликає скорочення м’яза, що він є причиною скорочення, я не проти. Можете уявляти це так. Але в голові ви повинні розуміти, що він не є прямим каталізатором скорочення, як у нейром’язовому синапсі скелетних м’язів. Він лише налаштовує темп спонтанної ритмічності, тобто визначає, наскільки близько до порога перебуває мембранний потенціал.
Я розумію, що це досить заплутано. Цю ідею потрібно добре засвоїти, ретельно обміркувати. Прокрутіть увесь процес у голові кілька разів, і тоді побачите, наскільки все стане зрозумілішим.
Далі я хотів би поговорити про розташування гладких м’язів. Вони є в таких місцях, як кровоносні судини, дихальні шляхи, де гладкі м’язи звужують або розширюють діаметр трубки. Якщо трубчасті структури, такі як кровоносні судини чи дихальні шляхи, містять у своїх стінках гладкі м’язи, то, коли гладка м’яза розслабляється, дихальні шляхи або судини розширюються. Якщо гладка м’яза скорочується, це призводить до звуження кровоносної судини чи повітроносних шляхів. Таким чином, ми можемо контролювати діаметр цих структур.
У всіх випадках гладка м’яза діє незалежно. Кожна гладка м’яза в судині має свій темп спонтанної ритмічності і, реагуючи на нейромедіатори, може мати швидший або повільніший темп. Подумайте про це: якщо у нас дуже повільний темп спонтанної ритмічності, настільки повільний, що клітина майже не скорочується, то гладка м’яза розслабиться, і трубка розшириться. Якщо у клітини високий темп спонтанної ритмічності, вона часто скорочується, і трубка звужується.
У інших місцях, наприклад, у кишечнику чи сечоводі, якими сеча стікає з сечового міхура, ми не просто змінюємо діаметр цих трубок. Уявіть кишечник. Ви бачили кишечник? Ходили на полювання на оленя, розкривали його і дивилися на його кишки, поки він ще теплий? Або, можливо, потрошили курку чи дивилися відео хірургічних операцій? Якщо розкрити когось і подивитися на кишечник, що він робить? Він здійснює червеподібні рухи. Ви чітко бачите ці рухи, що переміщуються. Як ми називаємо такі поступальні скорочення? Яку активність ми там бачимо? Як ми описуємо цей рух? Яке слово ми використовуємо, описуючи роботу кишечника? Що ви бачите, коли спостерігаєте за кишковим трактом? Ви бачите поступальний рух, і цей рух називається перистальтикою.
Ми спостерігаємо цю перистальтику, це хвилеподібне рух у кишечнику чи сечоводах, що переносять сечу з сечового міхура. Чи можете уявити, як виглядає перистальтика? Ось у нас є трубка, і гладкий м’яз в цій трубці скорочується, потім скорочується наступний гладкий м’яа, потім ще один, і так далі. Це було б схоже, якби я щільно обхопив трубку пальцями, стиснув її і провів пальцем по всій довжині трубки. Звісно, у нашому випадку немає зовнішнього тиску, лише скорочення гладких м’язів — одних, потім сусідніх, і так далі.
У місцях, де є перистальтичний рух, потенціал дії переходить від клітини до клітини. Таким чином створюється хвиля скорочень, рухома хвиля скорочень, тому що потенціал дії передається безпосередньо від клітини до клітини. Чи можете уявити цю картину? Ці клітини зчеплені разом досить дивним чином, і було б не зайвим описати природу цього зчеплення, але зробимо це іншим разом.
Наш час вичерпано. Не забувайте, що у вас тест у п’ятницю, тож готуйтеся. Побачимося тоді.